|
Introduction |
Isurus retroflexus (AGASSIZ 1843) was long thought to be present in the Lee Creek fauna, but to many collectors' surprise, it was not included by Purdy et al (2001:119); they deemed "I. retroflexa a nomen dubium", and included teeth of this design with Isurus hastalis (AGASSIZ 1843). Agassiz had erected Oxyrhina retroflexa based on a single, damaged, mako-like posterior tooth from the tertiary (location and stratigraphic position unknown). He noted the thin cusp, robust root and lingually directed curvature of the crown differentiated it from Oxyrhina xiphodon AGASSIZ 1843 (=Cosmopolitodus hastalis broad-form).
With such provenance, it would be easy to reject this taxon. However, René Kindlimann has provided information that sheds light on the original specimen likely from the Vienna basin, In addition, there is a published record for these thin-cusped makos; Cappetta (1987:96) included seven Mio-Pliocene citations. We maintain that this species is not only in the fauna, but present in sufficient quantities to reconstruct the tooth-set.
Materials & Methods
The Young collection includes many (900+) specimens from Lee Creek reject that have compressed cusps and a basio-lingual margin that extends beyond the root. From this collection, the junior author culled worn and damaged teeth, leaving approximately 500 specimens available (data-set). Representative examples (est. 175) of various tooth-designs / file positions became the basis for analysis (study-set)1. After the dentition-design was established, the data-set would be revisited for specific examples (size, coloration, etc.).
The vast majority of the mine's reject material originates from the Pungo River Formation (Early Miocene), however it may include contaminants from higher horizons or reworked material from lower. The quantity and condition of the study material strongly argues that it is largely Pungo River in origin.
In addition to various mako jaws, the authors had access to three tooth-sets: an Isurus oxyrinchus RAFINESQUE 1809 and two from the Hubbell collection (I. paucus GUITART-MANDAY 1966 and the "convergence" specimens.2 Rather than introduce the uncertainties raised by the "convergence" mako, only the other two were employed (2.2m Longfin and 1.9m Shortfin) for tooth-design purposes.
Each of the study-set teeth was digitally scanned from a labial perspective at 600dpi. Two forms of teeth were noted, broad and narrow-forms; in addition, many of the narrow-form teeth bore lateral cusplets. It was then deemed necessary to address each tooth-form separately and reconstruct each as an artificial tooth-set.3
Constructing each tooth-set followed a similar scenario. The labial images (a 2-dimensional evaluation) were digitally arranged using a generic lamnid rule-set4:
Following these propensities, each tooth-set would be hypothesized. These arrangements would then be double checked by evaluating the lateral perspective of the teeth. There is a general consistency of cusp curvature that should be reflected in these teeth as well. Of the studied teeth, 114 were ultimately scanned from a distal perspective. The resulting tooth-sets were remarkably similar to the extant Longfin, Isurus paucus. The lower A2 & A3 files were initially difficult to confidently ascribe to a particular position. In this case, a comparison of basal perspectives between I. paucus and these specimens was employed.
Narrow versus Broad Form
When viewing this material, it was apparent that cusps vary in width for similar file positions. In addition, a weak cusplet(s) can sometimes be found on the shoulders. As noted above, the retroflexus dentition-design would best be understood by developing two tooth-sets. Four general forms of variations needed to be considered.
Ontogenetic variations. The most readily observable correlation is the tendency for the cusp to expand relative to the cusp height as the tooth becomes larger. Figure 1 illustrates this property employing the upper A1s from the study-set.
 |
| Figure 1. Ontogenetic & Individual variations in Upper A1s |
Chronologic variations. Teeth with lateral cusplets are not characteristic of modern Longfins (possibly excluding young juveniles). As noted above, some Pungo specimens bear one or more cusplets. This propensity is largely limited to lateral teeth, most common in the narrow-form variety and more common in smaller teeth. These last two trends suggest that teeth with cusplets are the more primitive morpho-condition of the taxa. Figure 2 provides examples of this condition.
 |
| Figure 2. Cusplet examples |
Morphologic variations. Although all these variations are morphological in nature, one particular feature stood out and was noted in only six specimens. In all other specimens, the cusp broadened smoothly and extended onto the root (unless cusplets were present). In the case of these six, the cusp remained more upright as it approached the root. If no cusplets were present (A1 & A2) the base of the crown was mesio-distally compressed. When present (A3, L2, Lx), cusplets were low and extended. All six specimens appear to have come from similarly-sized individuals and the preservation appears identical. Based on the significance of this morphology, these specimens were excluded from the reconstructed tooth-sets5. Figure 3 provides examples of this feature.
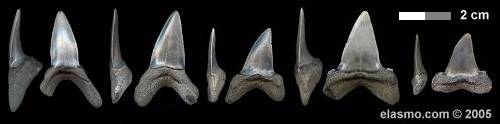 |
| Figure 3. I. cf retroflexus |
Individual variations. Excluding ontogenetic and chrono-based variations, teeth of similar size and file-position vary in design, which reflects individual variations. Figure one reveals these variations as well as ontogenetic.
Re-scaling Methodology
The similarity between I. retroflexus teeth and those of I. paucus is striking. For this reason a modern dentition was employed for tooth count and to assist in re-scaling. As discussed above, two tooth-sets (2.2m & 3.7m individuals) from the Hubbell collection were available, Since the identification of the larger specimen had been called into question (see Mako Dentition slides-how on website), only the smaller individual's teeth could be used at this time. The fossil teeth in the reconstructed broad-form dentition are slightly larger than those from the 3.7m specimen; therefore an ontogenetic bias may creep into the re-scaling, as the teeth of the 2.2m shark needed to be used.
Re-scaling was based on root width, not crown height. Comparisons will be based on the collective width of the anteriors and laterals 1-4. Crunching these numbers reflects an upper tooth-set remarkably consistent between paucus and the reconstructed retroflexus tooth-sets. As a group, the lowers are pretty close as a percentage of each group, although overall, the selected lower laterals appear to represent a larger individual than that of the uppers.
| Isurus paucus | Isurus retroflexus | |||
| Tooth Group | Root width | Root width | ||
| UA1-3 | 4.59 cm | 41.1% of total | 7.84 cm | 41.7% of total, 171.1% wider |
| UL1-4 | 6.58 cm | 58.9% of total | 10.96 cm | 58.3% of total, 166.6% wider |
| Uppers | 11.17 cm | 18.80 cm | ||
| LA1-3 | 4.66 cm | 46.3% of total | 7.67 cm | 42.5% of total, 164.6% wider |
| LL1-4 | 5.40 cm | 53.7% of total | 10.35 cm | 57.5% of total, 191.8% wider |
| Lowers | 10.06 cm | 90.1% of uppers | 18.02 cm | 95.8% of uppers |
Based on the similarity of the two upper tooth-sets, the upper retroflexus was deemed the baseline, and the lowers re-scaled accordingly: the total of the two lower retroflexus groups requires adjustment from 18.02 cm (95.8% of the uppers) to 16.94 cm (90.1% of the uppers).
Problematic are the retroflexus lower anteriors. Relative to root width, the crowns are lower than seen in I. paucus. This would suggest either a more extended anterior hollow or proportionately lower cusps; we have opted for the later.
With all this said, the retroflexus teeth were re-scaled based on the root width of corresponding I. paucus teeth. The lower laterals are most likely to be exposed to errors using this re-scaling basis. The scalebar will most accurately reflect the actual size of the upper teeth.
Summary
Accompanying are two slide-shows which include the tooth-set reconstructions (Broad & Narrow-form). Each compares well with the other, and although somewhat different to I paucus in tooth-design, the dentition-designs look similar to paucus (and distinctly different from I. oxyrinchus and Cosmopolitodus hastalis). The broad and narrow forms likely reflect a rapidly evolving morpho-species in the Early Miocene. Contrary to the Purdy et al (2001) arguments, there appear to be longfin makos in the Lee Creek fauna, and should be referred to as I. retroflexus at this time.
Footnotes
| 1. | Of the original study set, about 20 were eventually rejected as not representing I. retroflexus. These turned out to be thin-bladed variations of other isurids. |
| 2. | Refer to the "Isurus Dentition" slide-show for these tooth-sets and the problems associated with the "convergence" mako. |
| 3. | For the definition of an 'artificial tooth-set' as used on this website, see "Methods" page in "Reconstructed tooth-sets" slide-show. |
| 4. | The authors are uncertain of who might have been first to recognize various characteristics, although I suspect that root lobe divergence might be from Cunningham 2000. |
| 5. | Representing less than one-percent of the available teeth, there is no possibility at this time in reconstructing a corresponding tooth-set. |